  |
 |
 |
 |
 |
 |
 |
 |
 |  |
| ミトコンドリアは細胞のエネルギー代謝を支える「発電所」です。ミトコンドリアには、有機栄養素を酸素分子で酸化して炭酸ガスと水にまで変換し、その過程でATP(エネルギー)を産出する一連の酵素群が含まれています。特に、ATP合成に直接的に関わる呼吸鎖電子伝達酵素群はミトコンドリア内膜に埋め込まれています。 | |
|  |
哺乳類ミトコンドリアの呼吸鎖電子伝達系は、次の5つの酵素複合体から構成されています。○ NADH-ユビキノン酸化還元酵素(複合体-I)この内、電子伝達反応に共役してプロトンをポンプする酵素は、複合体-I、複合体-III、複合体-IVの3つの複合体です。ポンプされたプロトンによってミトコンドリア内膜を介して電気化学ポテンシャル勾配が形成され、複合体-V によるATP合成の駆動力となります。 複合体-II、複合体-III、複合体-IVおよび複合体-V(F1部分)の結晶構造解析が既に達成され、原子レベルで電子移動過程やプロトンポンプ機構が議論される時代に突入しました。 | |
|  |
|
ミトコンドリア複合体-Iは、大部分の還元力(電子)が呼吸鎖に流入する入り口に当たり、呼吸鎖によってポンプされる総プロトン数の半分を担っているエネルギー代謝にとって重要な酵素です。よく研究されているウシ心筋ミトコンドリア複合体-Iでは、45個の異なるサブユニットから構成される巨大な膜酵素(約1MDa)であることがわかっています。安定に単離・精製することは未だに容易なことではなく、呼吸鎖酵素中で最も研究の進展が遅れている酵素です。酵素内電子移動に関わるcofactorとしては、1個のフラビン(FMN)と8個のFe-Sクラスター群しかないため分光学的解析にも制約があり、このことも研究の進展が遅れている原因になっています。
| |
|
|
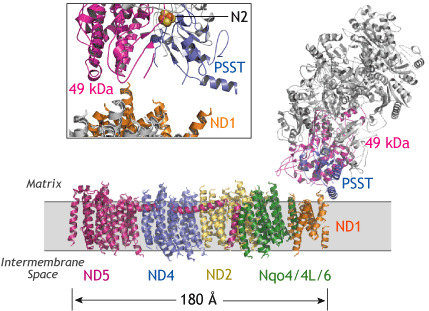
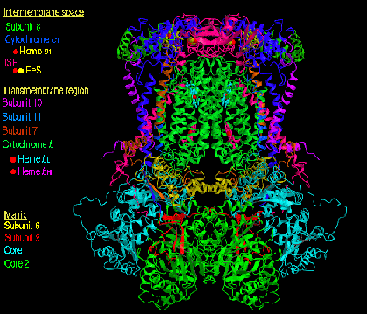
T. thermophilus複合体-Iの結晶構造 (Efremov et al. Nature 465, 441-445, 2010)
PDB entry: 3M9S, prepared by PyMOL v0.99. |
| 参考LINK (Scripps研究所、矢木研究室)→Complex I HomePage |
|  |
| ヒトでは複合体-Iの機能障害がパーキンソン症候群や遺伝性視神経委縮症などの重大な疾病(いわゆるミトコンドリア病)の原因となることがわかってきており、本酵素の基礎研究の大幅な進展が期待されています。また、最近の選択性に優れた農薬開発の有力な作用ターゲットとしても注目されています。 しかし、上記のように未だブラックボックスと言っても良い状況にあるのが複合体-Iです。「全くわかっていない。だから、我々の手法で挑戦したい!」というのが、複合体-Iを研究対象にしている理由です。 | |
|  |
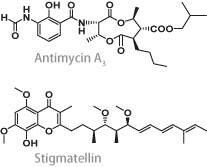
| 歴史的に眺めると、種々の呼吸鎖阻害剤が呼吸鎖酵素研究の発展に大きな貢献を果たしてきました。阻害剤の作用機構研究と呼吸鎖酵素の機能解明は両輪の関係で発展してきたわけです。例えば、複合体-IIIを考えた時、アンチマイシン(Qi部位阻害剤)とスティグマテリン(Qo部位阻害剤)がなければ、酵素内電子移動モデルとして今日では定説となった『Q-サイクルモデル』は確立しなかったでしょう。 | |
 | 
|
|
アンチマイシンとスティグマテリンの化学構造(左)、およびウシ心筋ミトコンドリア複合体-IIIの結晶構造(右)
Iwata et al., Science 281, 64-71, 1998 | |
|
我々は、呼吸鎖酵素に作用する独創的な生理活性プローブ分子をデザイン・有機合成し、世界にひとつしかないこれらの合成化合物を最大限に活用することにより呼吸鎖酵素、特にNADH-ユビキン酸化還元酵素(複合体-I)の機能解明を目指しています。 「モノを創り、モノを通して呼吸鎖酵素を探る!」 これが我々のスタイルです。 |
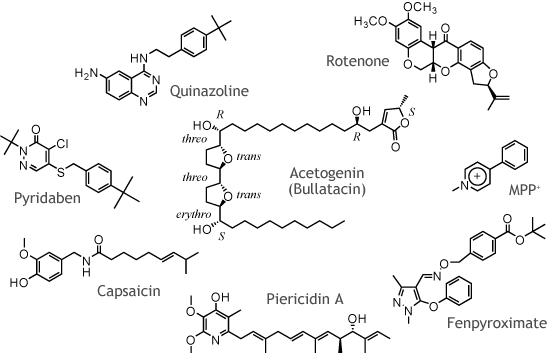 |
| 代表的な複合体-I阻害剤の化学構造 |
|  |
|
複合体-Iは、1電子あたり2個のプロトンをポンプすることがわかっています。他の酸化還元型プロトンポンプは、1電子あたり1個のプロトンしかポンプしません。この複合体-Iの特異なプロトンポンプ活性を説明するために、さまざまなモデルが考えられています。複合体-Iのメカニズムを知る上で、膜ドメインにユビキノン反応部位(= 阻害剤結合部位)が何カ所あるかということは重要な問題です。
そこで我々は、光親和性標識法によって種々の阻害剤の結合部位をアミノ酸残基レベルで明らかにしてきました。阻害剤は非常に多用な化学構造をとっているにも関わらず、PSST、49 kDaおよびND1サブユニットで構成される親水性ドメインと膜ドメインの境界領域(いわゆる”キノンポケット”)に結合することがわかりました。しかし、化合物の構造特性を反映して、結合様式は若干異 なっているようです。これは、複合体-IIIのQo部位阻害剤が、さらにサブタイプに分類されることを連想させます。 |
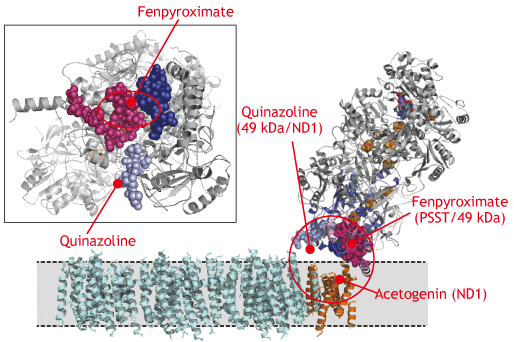 |
| 光親和性標識実験によって明らかにした種々の複合体-I阻害剤の結合部位 |
|  |
|
複合体-Iのプロトン輸送では、酵素の大きな構造変化が伴うことが示唆されています。
我々はこの動きを「1分子計測」技術で捉えることを目標に見据え、複合体-Iをピンポイ
ントで部位特異的に化学修飾する糸口を掴みました。
キノン結合ポケットを構成する49 kDaサブユニットのAsp160を様々な合成プローブで化学修飾します。 |
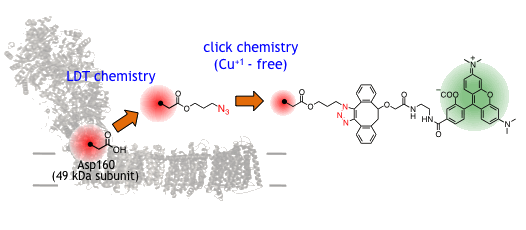 |
| ウシ心筋ミトコンドリア複合体-Iのピンポイント化学修飾 |
|  |
|
以上のようなエネルギー生産機関としてのミトコンドリア研究と平行して、”脂質”を基軸にして新たな角度からのアプローチを試みています。それは、細胞の自壊作用(アポトーシス)の制御機関としてのミトコンドリアの機能を解明することです。細胞に対する様々なストレスの多くはミトコンドリアに伝達され、その応答の結果としてアポトーシスが始まると考えられています。このミトコンドリアにおける応答において、重要な役割を担っているのがリン脂質の一種であるカルジオリピン(cardiolipin)です。
カルジオリピンはミトコンドリア内膜に局在するリン脂質で、呼吸鎖酵素やトランスポーターの安定化や機能発現に必須の成分です。アポトーシスの初動反応においては、ストレス信号である活性酸素によって過酸化を受けることで物理化学的性質が変化し、シトクロムcの放出や膜間孔の形成にも関与すると考えられています。しかし、これらの生理機能を制御するカルジオリピンの分子機構は全く明らかになっていません。そこで私たちは、カルジオリピン類のライブラリー構築とフィジカルプローブの創製を通して、カルジオリピンの機能解明を目指し研究を進めています。 | |
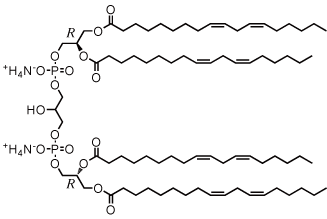 | |
| リノール酸で構成されるカルジオリピン類の構造 | |
|  |
|
|
|  |
|
更新中 | |
|  |
|
|
|